Nature:m6A调控miRNA初级体识别加工 | 案例解析
【字体: 大 中 小 】 时间:2018年05月24日 来源:联川生物
编辑推荐:
本文通过上述一系列严谨的实验,首次证明了m6A甲基化修饰是参与miRNA加工过程的一种新型调控方式。METTL3能够对pri-miRNA上GGAC motif的腺苷酸(A)进行m6A修饰。RNA甲基化修饰能够促进pri-miRNA被DGCR8特异性识别,从而成为miRNA成熟体加工过程中的一个十分重要的marker。
论文标题:N6-methyl-adenosine (m6A) marks primary microRNAs for processing
刊登日期:2015年03月
发表杂志:Nature
影响因子:40.1
研究机构:美国洛克菲勒大学
技术手段:m6A-seq,小RNA芯片,细胞敲低和过表达,qRT-PCR等
1.摘要
在miRNA生物合成的第一步是在微处理蛋白复合物的帮助下对初级microRNA(pri-miRNA)进行加工。微处理蛋白复合物由RNA结合蛋白DGCR8和III型核糖核酸酶Drosha组成。这个起始事件需要DGCR8识别pri-miRNA发夹上茎和侧翼单链RNA的连接处,并招募Drosha剪切双链RNA产生前体miRNA(pre-miRNA)。pri-miRNA的加工机制已经被阐释,但是目前并不知道DGCR8在众多具有二级结构的转录本中识别和结合pri-miRNA的机制。
本研究中,洛克菲勒大学Dr.Tavazoie教授的课题组发现在哺乳动物细胞中,METTL3会使pri-miRNA发生甲基化,标记的pri-miRNA可以被DGCR8的识别和加工。通过miRNA芯片分析发现,敲低METTL3会降低DGCR8与pri-miRNA的结合作用,引起成熟体miRNA表达量降低,通过未经加工pri-miRNA含量增加。体外实验证实m6A会促进pri-miRNA的加工。最后,功能验证实验揭示METTL3能够促使miRNA成熟。
本研究结果表明,m6A标记是一个关键的转录后修饰,会促进miRNA生物合成的起始。
2.METTL3直接介导pri-miRNA加工
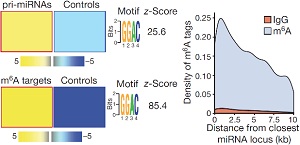
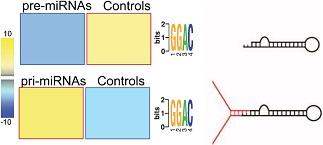
这篇文章的作者在先前的研究中已经证实,转录后的碱基修饰能够调控miRNA的加工过程。作者通过FIRE算法在pri-miRNA中找到了高丰度GGAC motif。这种motif与许多已发表的文献中METTL3识别的RGAC motif很相似。与很多pri-miRNA相反的是,这种motif并没有在miRNA前体序列(pre-miRNA)中富集。
接下来为了证实这种现象是否与METTL3酶相关,作者对乳腺癌细胞系MDA-MB-231使用了m6A-seq(也叫MeRIP-seq)测序,在m6A靶向的pri-miRNA上发现了许多GGAC motif。根据m6A-seq结果搜寻顺式调控元件发现METTL3 motif显著富集。
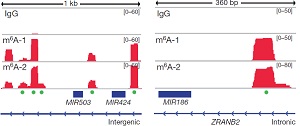
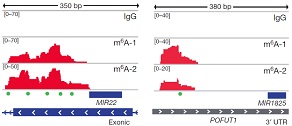
此外作者使用IGV软件发现,pri-miRNA上被reads覆盖程度较高的区域基本都是METTL3的motif并用绿色的小点标记了出来。
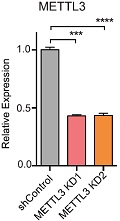
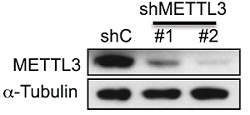
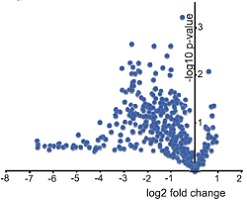
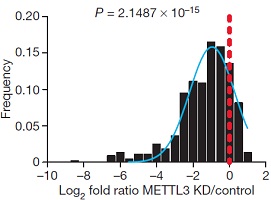
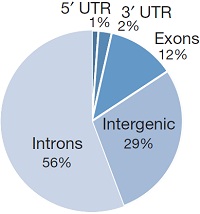
为了证实METTL3是否真的参与miRNA加工过程,作者在细胞系中使用两种短发卡RNA(short hairpin RNA)对METTL3进行干扰并用control shRNA进行对照,然后使用小RNA微流体芯片统计细胞中miRNA整体的表达情况。接下来作者发现miRNA成熟体整体表达量显著下调。此外,METTL3敲低后对miRNA在基因组不同位置上的影响呈现一定的多样性,内含子比例最高,接下来是基因间区。
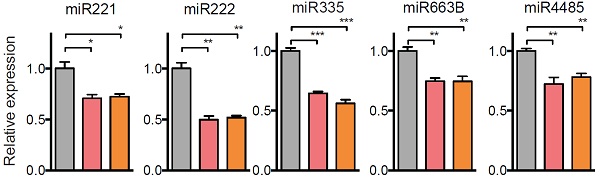
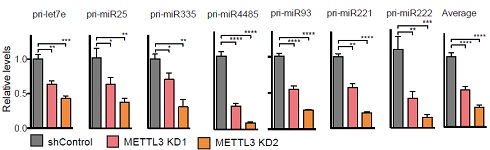
作者对几个关注的miRNA进行qPCR验证,发现受METTL3敲低影响,miRNA表达量显著下调。
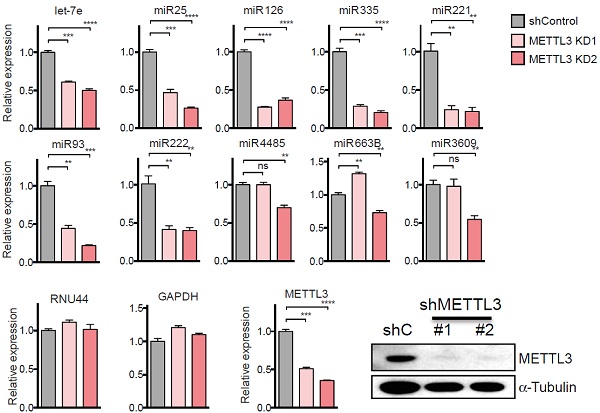
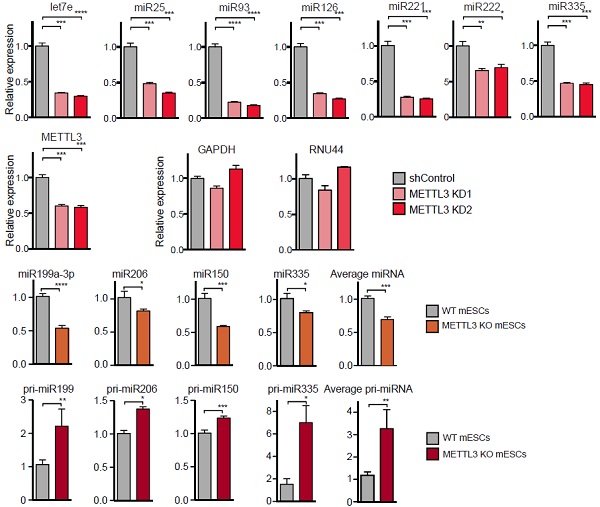
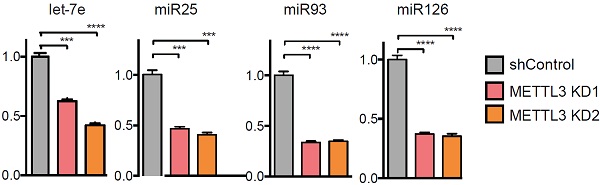
另外,METTL3敲低在多种哺乳动物的细胞系中都存在相似的情况,即miRNA表达量都显著下调。所有结果表明,METTL3无论在肿瘤细胞系还是常规细胞系中都对miRNA表达产生很大的影响。
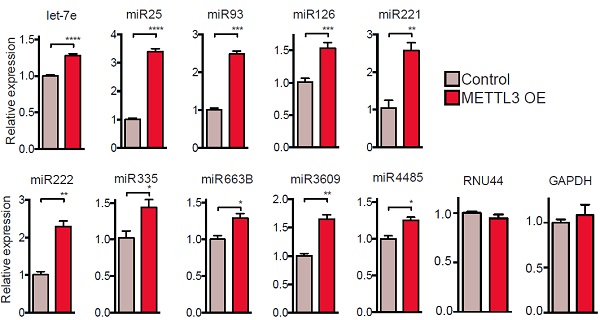
为了验证METTL3是否真正影响miRNA加工,作者在细胞系中转入METTL3过表达载体。结果表明METTL3过表达能够显著提高细胞系中miRNA的表达水平。
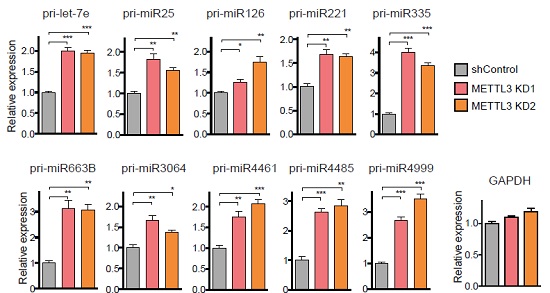
接下来作者发现METTL3敲低后,pri-miRNA表达量显著上升。这个结果表明pri-miRNA加工成pre-miRNA和miRNA成熟体需要METTL3来介导参与。
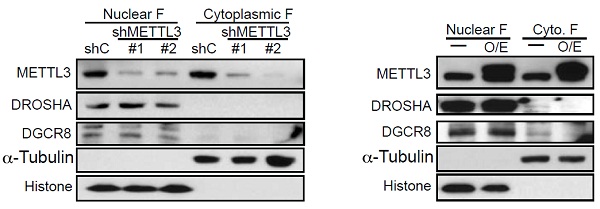
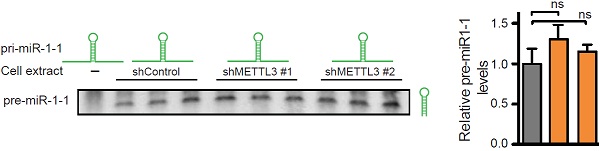
所以无论是METTL3的敲除敲低,还是过表达后改变了其表达量、细胞中亚定位或微处理蛋白复合物的催化活性。实验结果都表明METTL3直接参与pri-miRNA的加工过程。
3.METTL3直接参与pri-miRNA的m6A甲基化修饰
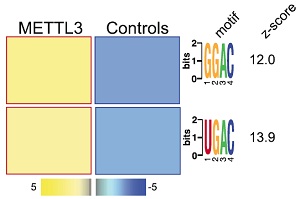
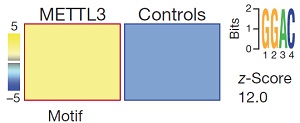
为了证实METTL3直接参与pri-miRNA的m6A甲基化修饰,作者使用METTL3抗体紫外交联法(CLIP-seq)进行验证。结果表明,与m6A-seq测序相似,METTL3 motif被显著富集。

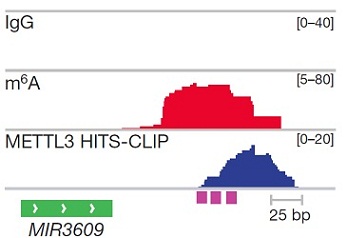
与m6A-seq测序结果相似,METTL3敲低/敲除后miRNA与METTL3 footprint存在极高的重叠率。
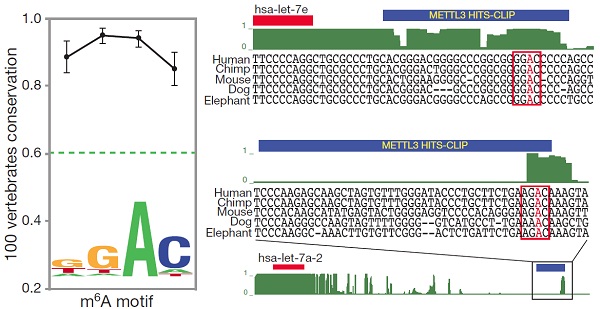
接下来作者采用Phastcons软件对100多种脊椎动物的miRNA以及METTL3 motif进行分析,结果表明METTL3在不同动物中都是极端保守的。
4.METTL3介导m6A修饰参与pri-miRNA加工
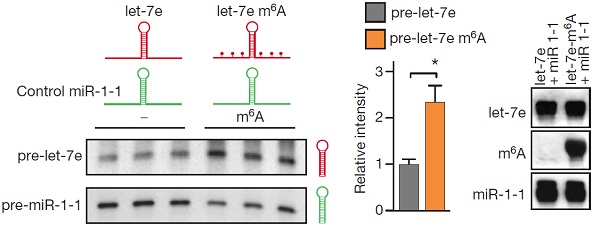
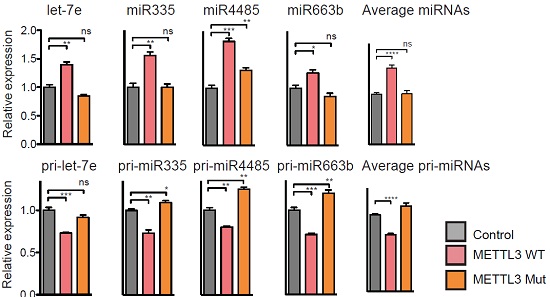
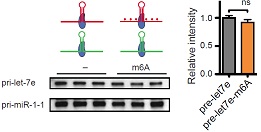
为了证明m6A修饰直接参与pri-miRNA的加工过程,作者获得了转染了DGCR8和DROSHA的HEK293细胞提取物。这些提取物被用于验证pri-miRNA是否含有m6A碱基修饰。通过Northern blot实验发现,发生甲基化的pri-let-7比未发生甲基化修饰的pri-let-7通过微处理蛋白复合物作用后产生pre-let-7的效率会更高。这个实验说明m6A甲基化修饰对于pri-miRNA加工成pre-miRNA这一过程至关重要。
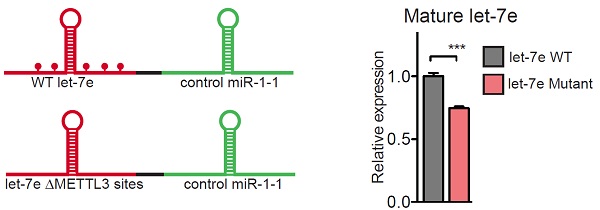
接下来,作者对pri-miRNA上容易发生m6A修饰的腺嘌呤苷酸(位于METTL3 motif中的A)进行点突变。已知pri-let-7e上一共含有五个腺苷酸(A),作者分别将五个A全部进行点突变。实验结果表明,这些被突变的METTL3 motif(也叫作腺嘌呤苷酸A)能够显著降低pri-miRNA加工成熟体miRNA的效率。
5.METTL3和DGCR8形成复合物参与pri-miRNA加工
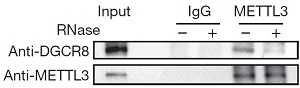
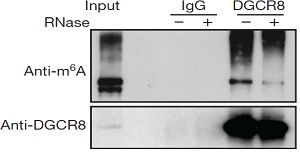
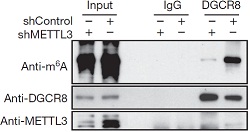
6.DGCR8特异性识别m6A甲基化pri-miRNA
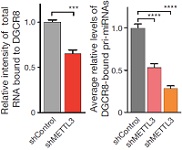
mRNA非常容易形成类似于pri-miRNA一样的二级结构。为了证实DGCR8是否真正能够特异性识别pri-miRNA而不是mRNA二级结构这个猜想,作者想要通过降低pri-miRNA的m6A甲基化水平,从而验证DGCR8蛋白结合的RNA总量是否所有下降。接下来,作者对细胞中的METTL3进行了敲除,并将所有的RNA进行放射性标记。DGCR8免疫共沉淀实验,METTL3敲除后DGCR8结合的RNA总量和pri-miRNA总量都显著降低(放射性信号减弱)。这些实验结果证实,发生m6A甲基化的pri-miRNA能够被DGCR8特异性识别,并影响后续成熟体miRNA的加工。
7.总结
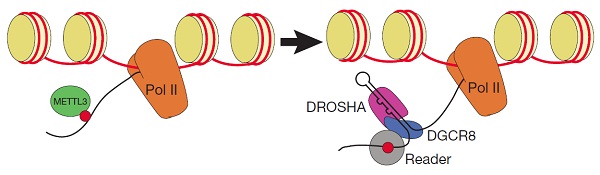
本文通过上述一系列严谨的实验,首次证明了m6A甲基化修饰是参与miRNA加工过程的一种新型调控方式。METTL3能够对pri-miRNA上GGAC motif的腺苷酸(A)进行m6A修饰。RNA甲基化修饰能够促进pri-miRNA被DGCR8特异性识别,从而成为miRNA成熟体加工过程中的一个十分重要的marker。所以作者认为m6A是一种十分重要的RNA标记,这种标记能够介导激活下游的微处理蛋白复合物,从而启动pri-miRNA向pre-miRNA和成熟体miRNA等一系列加工过程。因此m6A这种特殊的标记在细胞核中具有重要的作用,即允许微处理蛋白复合物识别具有特定二级结构且发生甲基化的pri-miRNA而不是其他一些底物如具有二级结构的mRNA等。此外,作者也推测METTL3的异常表达可能会引起肿瘤中miRNA的异常高表达。




